|

01动植物繁殖博弈中的代价与平衡
在自然界,约90%的开花植物依赖动物传粉。绝大多数传粉关系是简单的互利交换:你取食花蜜,我完成授粉。但有一类关系更为特殊,也更具戏剧性—育幼传粉。在这种传粉模式下,植物将子房提供给传粉者作为后代发育的场所,传粉者则在访花时帮助授粉。听起来公平?然而这里藏着一个根本性的矛盾:传粉昆虫的幼虫孵化后,必须以植物的种子为食。植物“雇佣”昆虫传粉,代价是牺牲一部分后代。如果这笔交易失衡,植物将血本无归。那么,这种微妙的共生关系,为何能在漫长的进化过程中维持稳定?中国科学院昆明植物研究所陈高团队的一项研究,在水龙与其专性传粉象鼻虫之间,找到了一个新的答案。这场交易最终的“仲裁者”,既不是植物,也不是昆虫,而是“水”。
02花朵未开,虫卵已“种下”
水龙(Ludwigia adscendens)是一种柳叶菜科丁香蓼属多年生浮水草本植物。花期5-9月,其花朵在水面以上10—20厘米处绽放,单花寿命仅一日。野外观察发现,一种体型很小的象鼻虫(Nanophyes proles)是它的主要传粉者。象鼻虫不仅在花上取食花蜜,频繁活动于花药与柱头之间,还在花上交配,花朵同时充当其觅食、交配和繁殖的场所。开花前2~3天,花尚处于花苞期,雌虫便用坚硬的喙刺穿子房壁,随后将卵产入子房中。一个果荚内可寄生1至3枚卵。从幼虫孵化、化蛹到成虫羽化并钻出果荚,整个发育过程均在果荚内完成。也就是说,在花朵开放之前,象鼻虫的后代已经被安置在未来的果实之中。
03被象鼻虫“操控”的植物
寄生行为对水龙的花和果实形态产生了显著影响。被寄生的花朵,其花柱长度显著缩短,而花丝长度不变。这一变化促进了自花授粉。即使在缺乏传粉者的情况下,植物仍能通过自交产生种子。这既为植物提供了繁殖保障,也确保了果荚内有种子可供幼虫发育所需。此外,被寄生的果荚形态变得更短、更粗。这种改变可为幼虫提供更宽敞的发育空间。

图1(a-c) 水龙的植株、生境,象鼻虫在花上取食和交配;(d-e) 象鼻虫在花上产卵;(f-g) 被寄生和未被寄生的花、果荚的对比;(h-k) 果荚内发育的幼虫和蛹及成虫逃逸的孔;(i-n)沉水的果荚及淹死的幼虫和蛹。
04三个幼虫,吃掉95%的种子
然而,任何合作都有底线。这项交易是否划算,关键要看代价。两年的调查数据显示,水龙果荚的寄生率高达52%至56%。也就是说,超过一半的果荚里都养着“别人家的孩子”。种子消耗量与果荚内的幼虫数量呈正相关:一个幼虫大约吃掉33%的种子;两个幼虫消耗过半;一旦幼虫数量增加到三个,种子损失超过95%。当果荚内同时寄生多只幼虫的频率升高时,植物所承受的代价将显著增加,互惠关系面临崩溃的风险。
05果实沉水的调节功能:一种物理“制裁”机制
平衡植物与传粉者博弈的关键在于水。水龙的果荚在花后3至4周主动沉入水中。在自然条件下,72.2%的象鼻虫能够在果荚沉水前完成羽化并逃逸,而27.8%的个体因未能及时羽化而被淹死。在被淹死的个体中,64%处于幼虫阶段。象鼻虫被淹死的果荚中留存了更多的种子。水淹通过杀死部分未及时发育成熟的幼虫,间接减少了幼虫对种子的总体消耗。水淹起到了类似陆生植物“主动制裁”的功能,但实现路径完全不同:不需要化学信号识别,不需要主动落果决策,仅仅是生活史时序与水生物理环境的耦合,便在长期互作中持续维持着共生关系的平衡。
06从陆地到水域,从主动到被动
该研究拓展了育幼传粉系统的生态边界。此前已知的育幼传粉系统均存在于陆生植物(如榕树与榕小蜂)或沼泽植物中。该研究首次在真正的漂浮水生植物中证实了该系统的存在,将此类共生关系从陆地拓展至水生生态系统。更重要的是,该研究揭示了一种非生物的冲突调解机制。这并非任何一方精心“设计”的结果,而是植物和昆虫在长期博弈中形成的一种动态平衡和生存智慧。
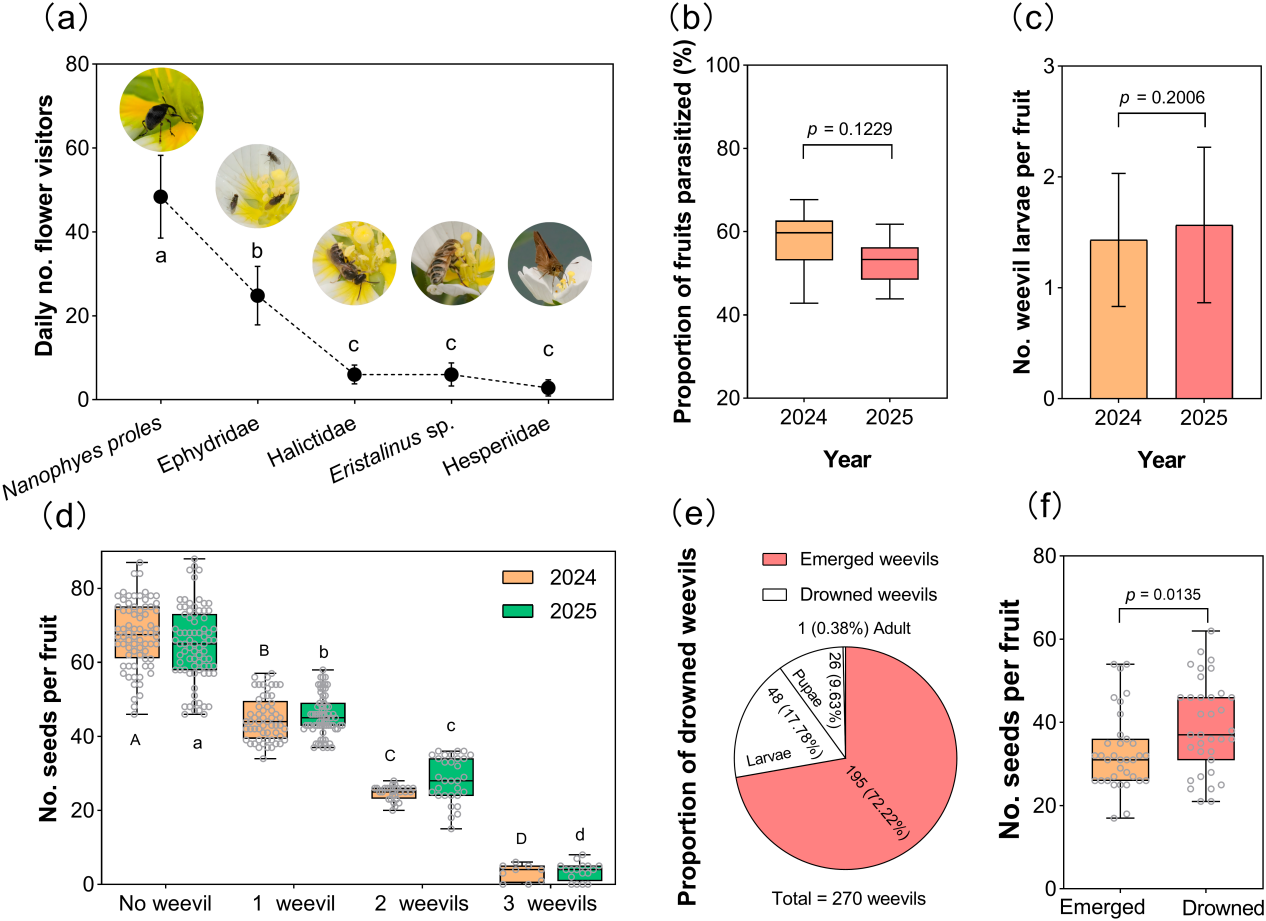
图2 (a)水龙传粉者的访花频率;(b-c)果荚寄生率及果荚内寄生的幼虫数量;(d)不同寄生程度消耗种子的情况;(e)象鼻虫羽化和淹死的比例以及发育阶段;(f)象鼻虫逃逸和淹死剩余种子数量
近日该研究以A novel nursery pollination mutualism between a water primrose and its fruit-parasitizing weevil为题发表于国际生态学经典期刊Ecology。中国科学院昆明植物研究所的博士后吴事茂为论文第一作者,陈高研究员为通讯作者。该研究得到了云南省科技计划项目(202401AT070194)、云南省科技厅项目(202303AC100011)及国家自然科学基金项目(32500191)的资助。
文章链接
|

