|
多倍化或全基因组复制(WGD)是植物演化的重要驱动力。2011年,研究人员基于基因树定年分析推断种子植物和被子植物祖先分别发生了ζ-WGD(约3.19亿年前)和ε-WGD(约1.92亿年前)事件(Nature 473: 97–100)。然而这些WGD事件的真实性长期存疑。2026年,相关学者利用剂量敏感基因的保留模式证实了ζ-WGD事件但否定了ε-WGD,使这一争议再度成为焦点(Science Advances 12: eaea9797)。
研究团队首先采用直系共线性分析系统比较了裸子植物和被子植物代表性基因组。银杏的染色体结构被认为与祖先裸子植物核型高度相似(Advanced Science 12: 2411098),为提供高质量的裸子植物参考基因组,团队对银杏(Ginkgo biloba)基因组进行了重新注释,重新注释后BUSCO完整性从63.5%大幅提升至93.1%。
尽管存在共线性碎片化,银杏与无油樟(Amborella trichopoda)之间的直系共线性呈现出清晰且一致的1:2对应关系(图1A)。在4520个滑动基因窗口中,银杏的一个基因组区域对应无油樟的两个不同直系同源区域(图1A)。由于无油樟自与现存其他被子植物的共同祖先分化以来未经历WGD,因此全基因组范围的1:2共线性模式为被子植物祖先在与裸子植物分歧后经历的一次四倍化事件(即对应ε-WGD)提供了强有力的证据。这一直系同源信号在其他被子植物中也保持稳定:未经历独立WGD的物种(如马兜铃属Aristolochia)保留1:2模式,而经历后续WGD的谱系表现出更高的比例,与其独立的多倍化历史一致(图1B–C)。
与之形成鲜明对比的是,银杏与其他裸子植物基因组之间显示1:1的共线性比例(图1B–C),而这些裸子植物与被子植物比较时也普遍表现出1:2或更高的比例(图1D)。这一对比模式有力地表明ε-WGD发生于被子植物共同祖先。此外,在无油樟基因组中可检测到古老的旁系同源共线性(图1E),尽管其重复保留率较低,但也为ε-WGD事件提供了进一步的基因组结构证据。
为进一步验证ε-WGD,研究将代表性被子植物的共线性区块划分为两个亚基因组,并进行了系统发育分析。结果表明两个亚基因组为姐妹谱系,91.9%的单基因树与该拓扑一致(图1F)。尤为关键的是,这两个亚基因组的分歧发生在银杏与被子植物谱系分化之后、但在现存被子植物分化之前(图1F)。亚基因组系统发育与直系共线性模式高度吻合,强烈支持被子植物祖先基因组的加倍源于一次共享的被子植物WGD,确立了ε-WGD作为被子植物演化的奠基性事件(图1G)。
该研究成果以Orthologous synteny provides robust structural evidence for the ancestral angiosperm ε-WGD为题发表于植物学顶刊Molecular Plant。中国科学院昆明植物研究所博士研究生张仁纲为论文第一作者,马永鹏研究员为通讯作者。捷克马萨里克大学Martin Lysak教授、昆明植物研究所博士研究生尚鸿运、中国科学院植物研究所焦远年研究员参与了该研究。该研究得到国家重点研发计划(2022YFF1301700)、中国科学院昆明植物研究所战略先导科技专项(KIBXD202401)等基金的资助。
文章链接
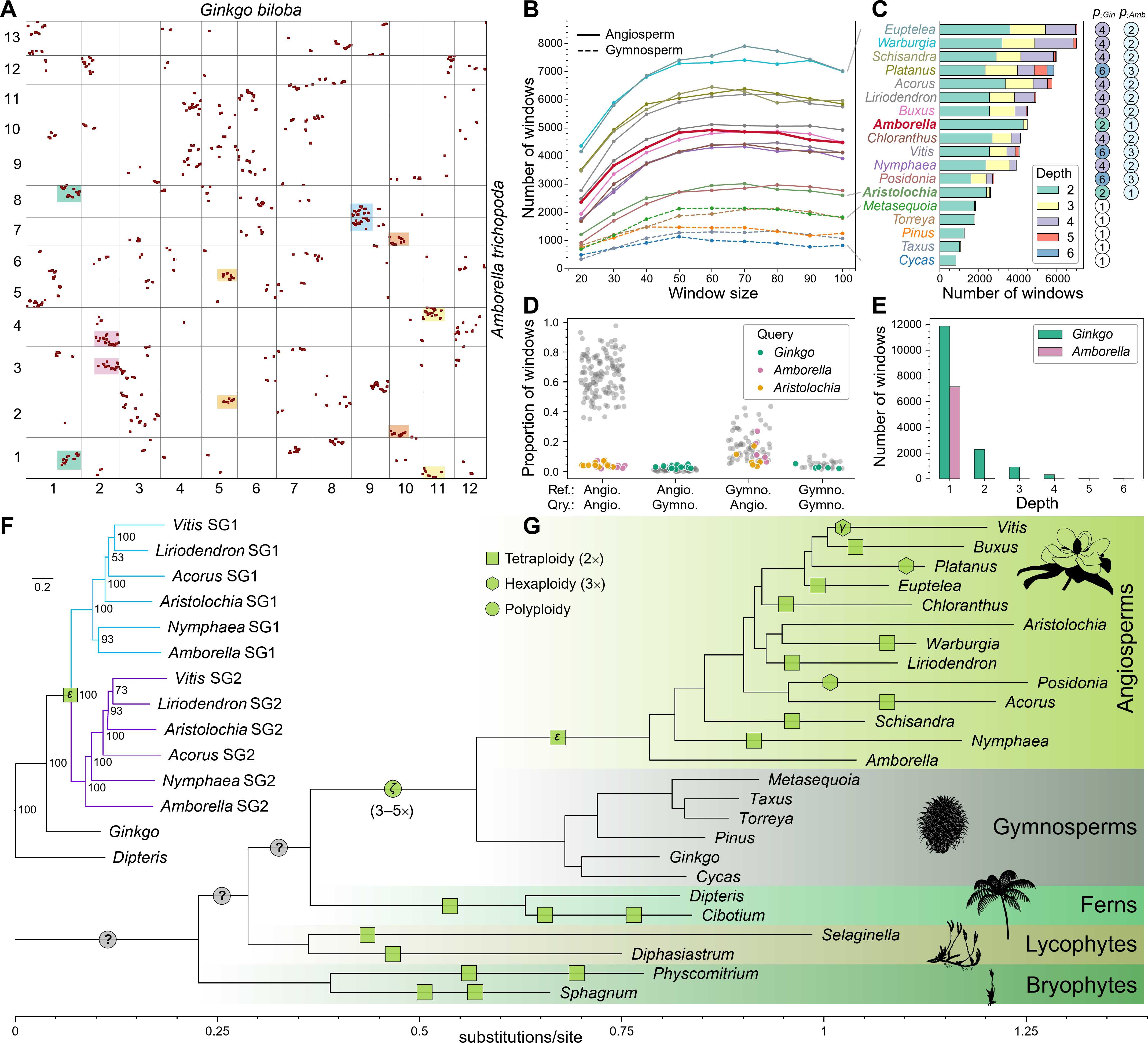 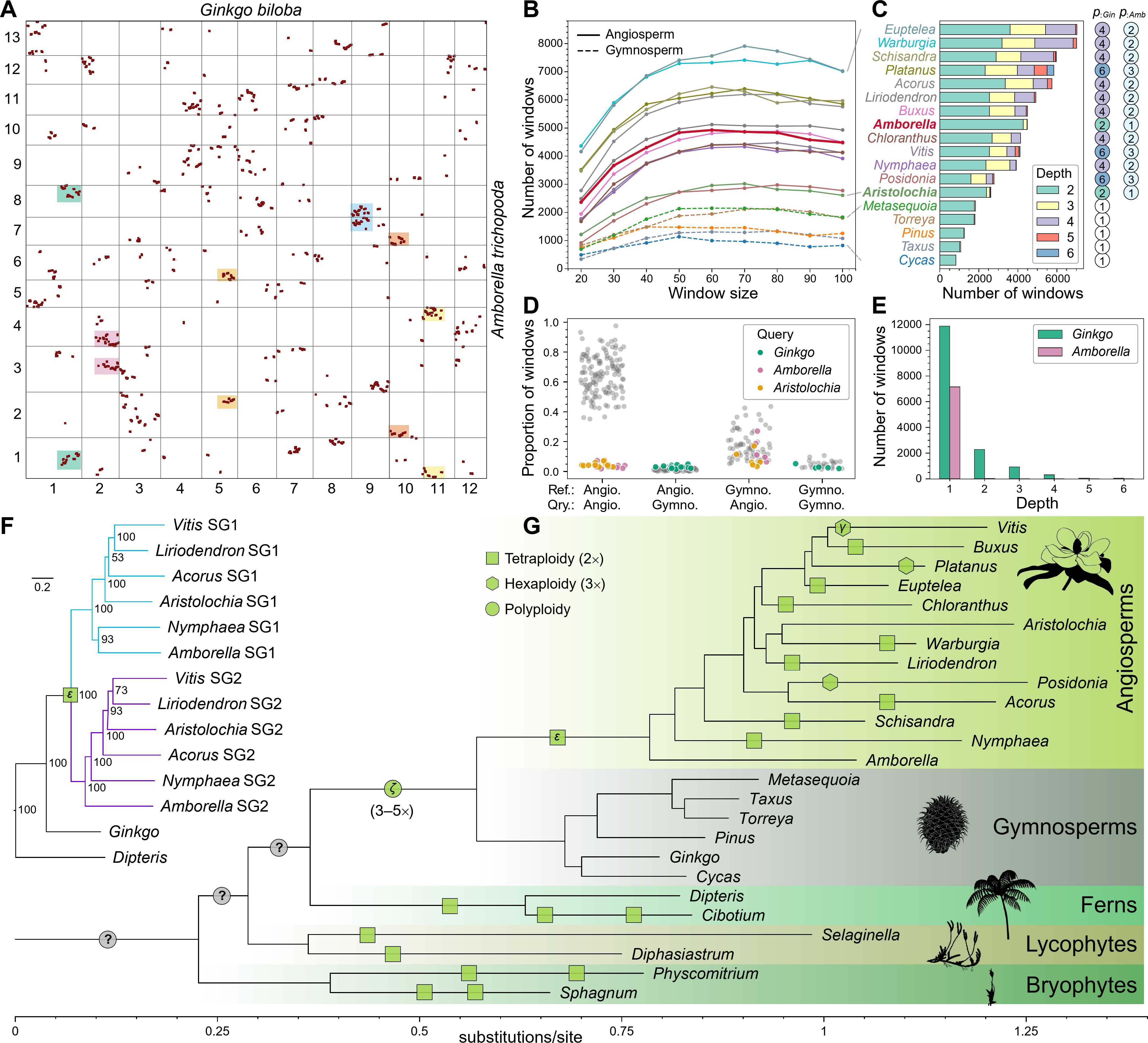
图1 支持被子植物祖先ε-WGD的直系共线性与系统发育证据
(A)银杏与无油樟基因组的直系共线性散点图,坐标轴数字为两基因组染色体编号,每个点代表一对共线性基因,彩色阴影标注多处1:2对应的示例。(B)以银杏基因组作为参考,20—100个基因滑窗下深度≥2窗口的丰度。(C)50个基因窗下各物种相对于银杏的共线性深度组成,右侧圆圈标注基于共线性深度模式推断的相对于银杏和无油樟的倍性。(D)两两基因组比较中深度≥2窗口的比例,每个基因组依次作为参考计算其他基因组的共线性深度。(E)银杏和无油樟基因组内部的旁系同源共线性深度分布。(F)基于234个直系同源基因集构建的被子植物两个亚基因组(SG1, SG2)与银杏的系统发育关系。(G)陆地植物系统发育框架中推断的WGD事件位置,不同形状代表不同倍性的多倍化事件,绿色填充为共线性证据支持的事件,灰色填充为本研究基于剂量敏感重复基因保留模式的推测。
|

