|
自然界随处可见形态各异的植物叶片,分为单叶和复叶两种基本类型;其中,由多个小叶排列组成的复叶(如含羞草、豌豆的叶片)以其精巧复杂的结构尤为引人注目。复叶是如何发育形成的?其形态多样性背后蕴含着怎样的进化智慧?近日,中国科学院西双版纳热带植物园(以下简称“版纳植物园”)陈江华研究团队在世界知名植物学期刊Current Opinion in Plant Biology上发表了题为“Multilayered regulatory control of compound leaf development”的综述文章,系统梳理了复叶发育研究领域的最新进展,特别聚焦于KNOXI、LFY/FLO和CUC/NAM等核心遗传模块,阐述了它们如何通过与激素信号和其它核心调控因子相互作用来塑造丰富的复叶形态。
复叶发育起始于茎顶端分生组织(SAM)周缘区的简单钉状(peg-like)原基。最早期的复叶原基,其侧缘分生区(Marginal Blastozone)维持短暂的分生组织活性,是小叶原基起始的干细胞生物学基础。经典的KNOXI、LFY/FLO和CUC/NAM等核心遗传模块在维持、调控侧缘分生区活性功能上扮演了关键角色。
在大多数非豆科复叶植物(如番茄和碎米荠)中,KNOXI基因在SAM和早期复叶原基中表达,维持复叶侧缘分生区的干细胞活性来促进小叶原基形成。而在豆科IRLC植物(如蒺藜苜蓿和豌豆),花分生组织特征基因LFY/FLO替代了KNOXI功能,调控小叶原基的起始。这些模块与激素信号(如生长素、细胞分裂素)紧密互作:生长素通过PIN1介导的极性运输在小叶起始位点形成浓度峰值,而KNOXI或LFY/FLO与生长素信号途径形成精细的互反馈调控,共同决定小叶原基的有序发生,并影响其数量和排列方式。同时,CUC/NAM转录因子在侧生器官边界(包括小叶边界)特异表达,确保小叶原基发育为独立单元。这些核心模块的相互作用构成复叶形态建成的基本框架。
复叶形态的多样性源于进化过程中核心遗传模块的“重新布线”。诸如基因复制(gene duplication)、新功能化(neofunctionalization)及顺式调控元件的改变(cis-regulatory changes)等机制,使保守模块在不同物种中产生出独特的作用和表型输出。例如,番茄中KNOXI与BLH蛋白的互作、碎米荠中RCO基因的演化,分别调控了小叶数目和精细的叶轴结构;IRLC豆科植物复叶中KNOXI功能的退化与LFY/FLO模块的招募来替代其功能等。最近的研究表明:一类C2H2锌指蛋白(PALM1/MPL1)作为关键的复叶原基侧缘分生活性负调控因子,与LFY/FLO形成保守的“拮抗”模块,来精确调控后者的活性强度与时空表达模式,并与生长素信号、CUC/NAM边界基因以及其它物种特异性因子(如豌豆的TENDRIL-LESS)等协同互作,共同决定了从苜蓿三出复叶到鹰嘴豆羽状复叶、乃至豌豆具卷须的特殊复叶等千姿百态的复叶形态建成。
该文由版纳植物园陈江华研究员和贺亮亮副研究员担任共同通讯作者,贺亮亮、杨丽玲博士和赵维月博士为的并列第一作者。研究得到国家自然科学基金、云岭学者等项目的资助与支持。
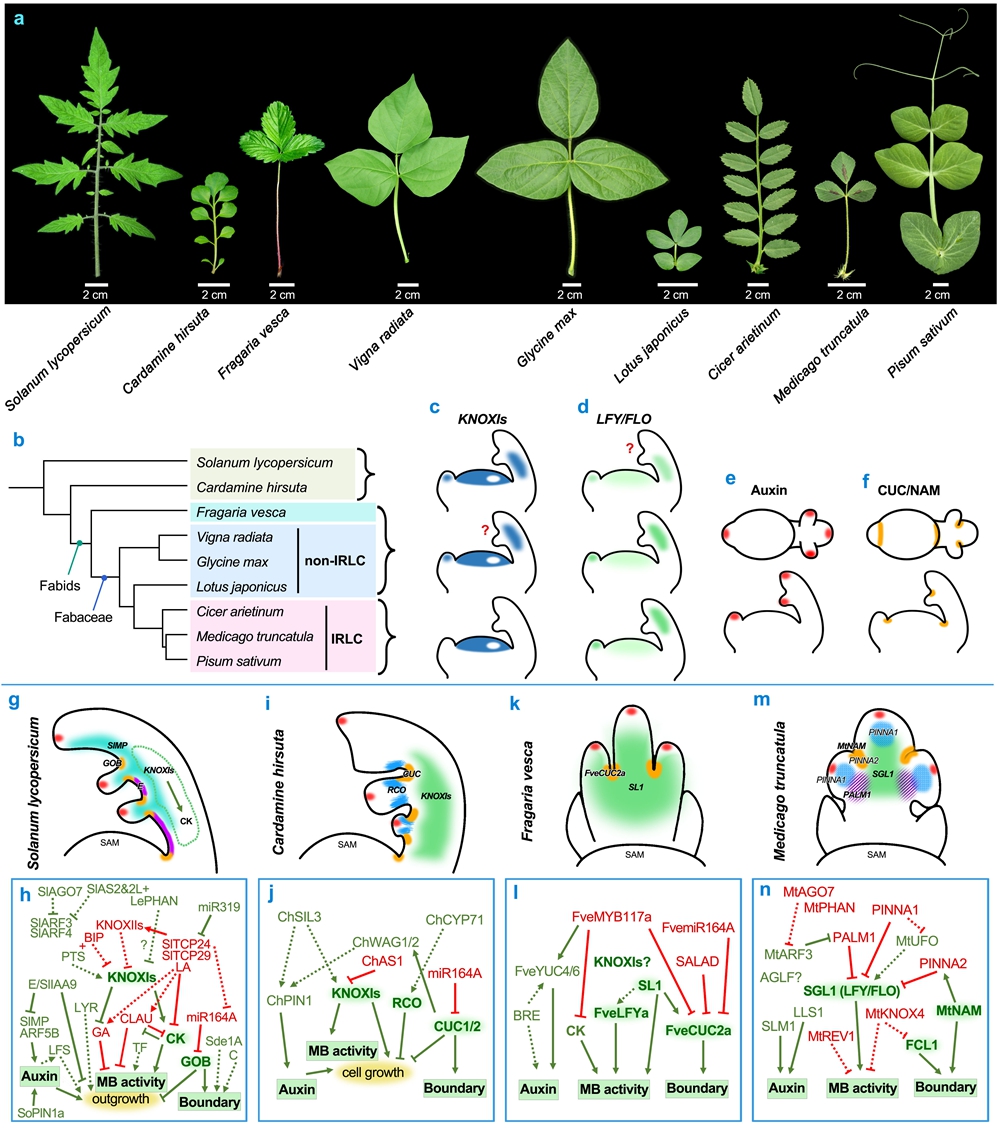
图1:复叶的多样形态及其发育调控网络。KNOXI、LFY/FLO和CUC/NAM等核心遗传模块,通过与其他调控因子及激素信号互作,共同塑造复叶形态的多样性。
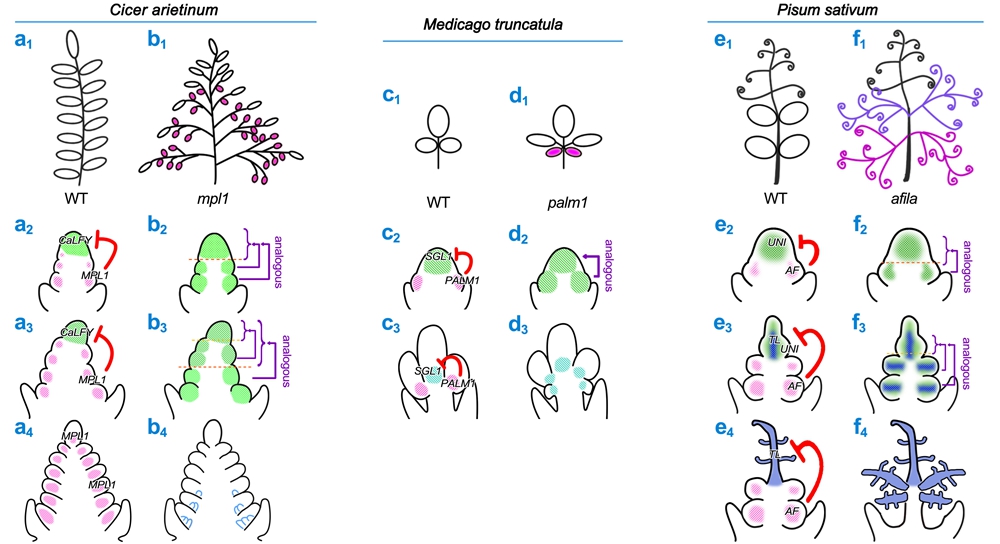
图2:由C2H2锌指蛋白PALM1/MPL1/AF和LFY/FLO构成的保守“拮抗”模块的时空特异性调控,是介导从苜蓿的三出复叶到鹰嘴豆的典型羽状复叶、豌豆的具有卷须的特殊复叶的形态建成的关键。
|

