|
花朵的千姿百态从何而来?科学家发现,传粉昆虫的“偏爱”是重要推手!被子植物为吸引传粉者,演化出精妙的花部结构。有趣的是,在亲缘关系较远的植物中,相似的传粉环境会让花朵“殊途同归”—形成趋同演化的形态特征。然而,同一性状趋同演化背后的基因密码是否相同是进化生物学的核心谜题之一。异型花柱(heterostyly)现象堪称植物花结构的精巧设计:同一个物种的群体中存在两种或三种花型,雌蕊和雄蕊高度呈互补排列(如高柱头配矮花药,矮柱头配高花药),这种花部结构的“交互异位”不但能够促进花粉精确传递,促进异株授粉,维持种群多样性,而且还能减少花粉干扰与避免近亲繁殖。异型花柱这类植物花部结构多态性现象至少在被子植物28个科中被发现,其中茜草科中发现有最多的属和种具有异型花柱的性系统,然而调控花柱长短相关基因及分子调控机制却仍然成谜。
中国科学院华南植物园罗世孝研究团队以茜草科玉叶金花属(Mussaenda)为研究对象,结合基因组从头组装、比较基因组、群体基因组、转录组和系统发育等手段,鉴定了控制玉叶金花属异型花柱发育的超基因(S-locus supergene)的结构及其起源演化规律。研究发现(1)控制玉叶金花属异型花柱发育的超基因由3个紧密连锁的基因(MuIAA, MuGA3ox, MuAPs)组成,且只存在于短柱花个体中,呈半合子结构;(2)由于半合子结构导致的重组抑制,S-locus 所在基因组区域比其两侧区域富集显著高比例的重复序列;(3)S-locus 超基因是通过基因的逐步复制并且插入到该区域所形成的, 其中MuIAA 的复制出现在茜草科分化之前,而MuGA3ox 则出现在茜草科物种分化之后;(4)超基因中名为MuIAA的生长素响应基因在短花柱的雌蕊和花筒部位高度活跃,是调控花柱长短的候选基因。该研究不仅证实异型花柱演化在分子上存在趋同机制(S-locus 的普遍半合子结构),更首次揭示生长素相关基因在调控花柱长短的核心作用。未来进一步的科学问题包括为什么某些植物类群更易演化出异型花柱?激素代谢通路如何影响这类精巧结构的存续?这些发现将为解析植物多样性起源打开新窗口。
相关研究成果以“Genomic evidence unveils the genetic architecture and evolution of the S-locus controlling heterostyly in Rubiaceae” 为题于近日发表于植物学期刊New Phytologist (《新植物学家》)(IF5year=10.3)上。华南植物园副研究员袁帅为论文第一作者,罗世孝研究员和张奠湘研究员为共同通讯作者。该研究得到国家自然科学基金(32370239)和华南植物园青年人才专项(QNXM-06)的资助。文章链接:https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.70311

图1. 三种玉叶金花属植物图片。下侧解剖图分别展示长柱花和短柱花结构。
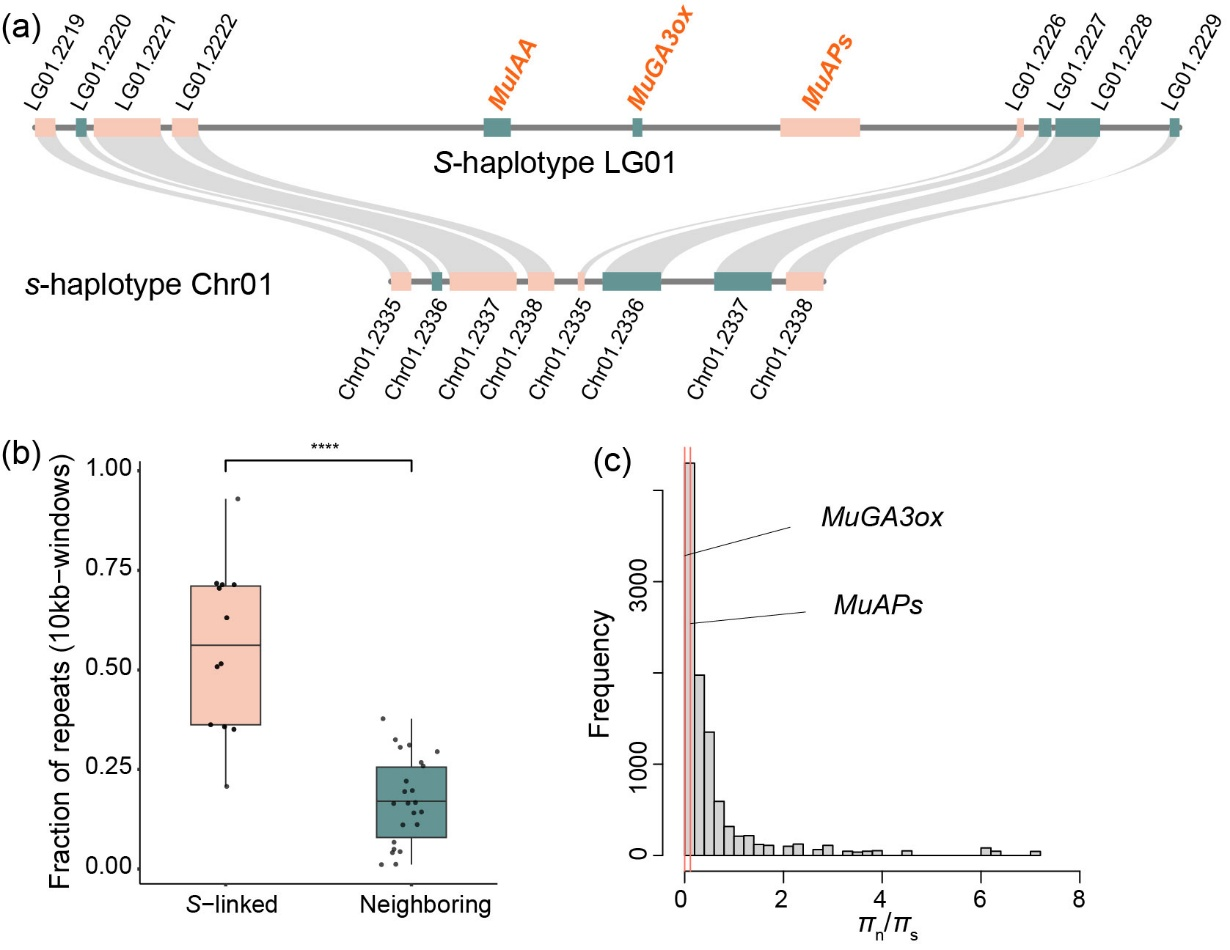
图2. 玉叶金花属S-locus 超基因的半合子结构及其分子进化规律。
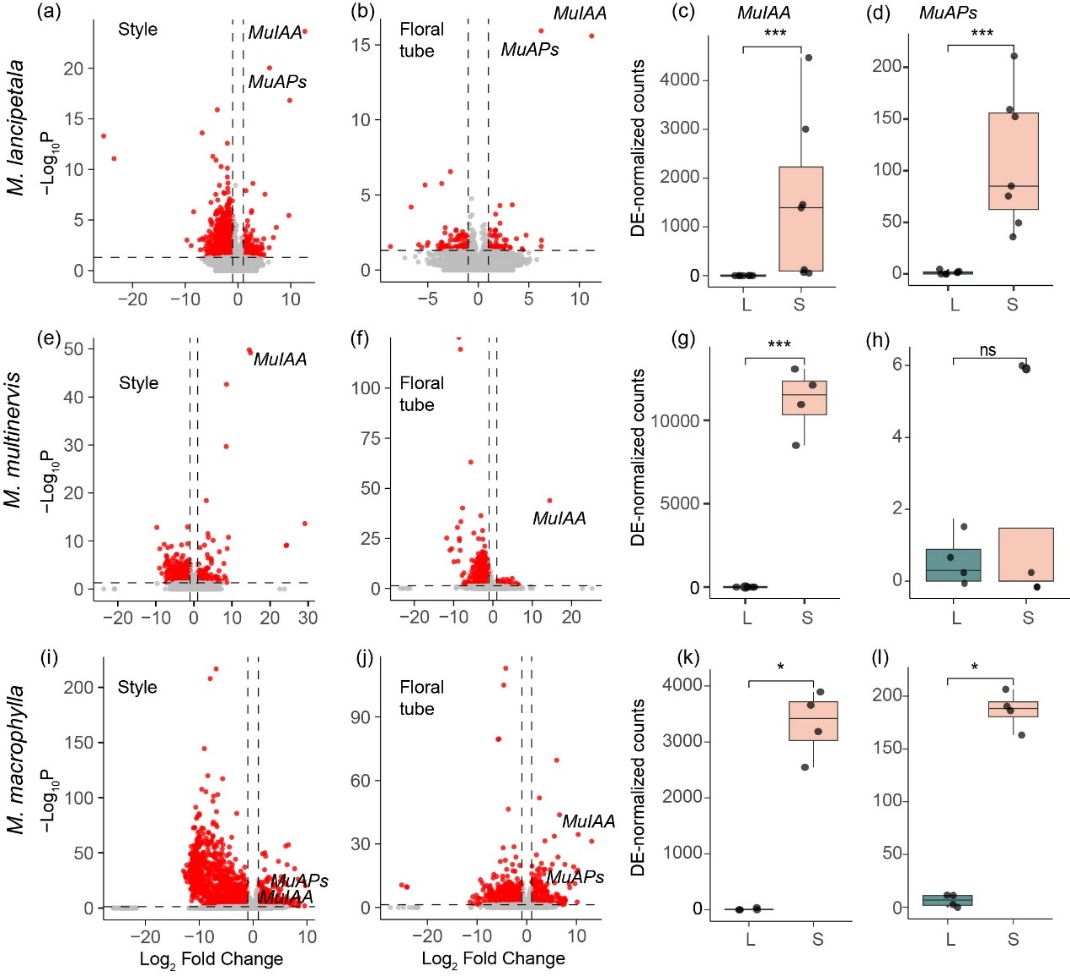
图3. 玉叶金花属长柱花和短柱花的花柱和花冠管的差异表达基因。S-locus 基因MuIAA 和MuAPs 在短柱花中显著高表达。
|

